|
|
||||||||
| Artículos | Casos | Libros | Apuntes | Otros | ||||
| |||||||||||||||||
|
Librería Biopsychology.org
La ecología es la ciencia que estudia el comportamiento de los seres vivos en todas sus facetas y dimensiones. Es, por tanto, una macro-ciencia, una ciencia general que contiene muchísimos campos específicos. Prueba de ello es este manual ciertamente extenso y completo. A diferencia de los enfoques de las ciencias del comportamiento humano tradicionales, la ecología trata el comportamiento desde un punto de vista estrictamente real y objetivo. Los ecólogos no tienen que salvar el prestigio ni el orgullo de ninguna especie y, por tanto, pueden dedicarse al estudio del comportamiento sin prejuicios morales. A lo largo del siglo XX, la ecología ha desarrollado un cuerpo de conocimientos suficientemente extenso para convertirse en el punto de referencia de todo aquel que quiera comprender el comportamiento humano real y verídico. La ecología trata fundamentalmente de las relaciones entre los seres vivos y las consecuencias en su supervivencia. La biopsicología es, pues, una subdisciplina de la ecología, en tanto que nuestro objetivo es el estudio de las relaciones humanas y su incidencia en la salud y el bienestar (nuestra supervivencia al fin y al cabo). Gracias a la ecología, disponemos de un amplio abanico de conocimientos y conceptos generales obtenidos del estudio de un gran número de especies distintas. Por supuesto, los seres humanos somos una especie distinta y única, como lo son todas las demás. Esto significa que no todo el trabajo está hecho. Partiendo de una base general proporcionada por la ecología, la biopsicología tiene un largo camino por recorrer investigando las formas particulares de relación social de nuestra especie. Así que, cualquiera que desee formarse en biopsicología debe estudiar a fondo ecología. No es una materia colateral ni adecuada, es imprescindible, necesaria, y casi suficiente. De hecho, la ecología es la antesala de la psicología. Querer hacer psicología sin pasar por la ecología es estar en un sitio equivocado, probablemente haciendo filosofía o cualquier otra cosa menos psicología. Este modo de enfocar la psicología sorprende a muchos debido a que se tiene una falsa imagen de la ecología por culpa del ecologismo. Al ser esta última una corriente política, tiene mucha más audiencia que la primera y es más conocida. Pero el ecologismo tiene que ver con la ecología lo mismo que la democracia cristiana tiene que ver con el cristianismo, es decir, nada de nada. Una es política y la otra es ciencia y, normalmente, ambas actividades se excluyen mutuamente. Tenemos el gusto de ofrecer al lector un espléndido manual de ecología con el que podemos asegurar que podrá comprender mucho mejor la naturaleza del comportamiento humano. Para el futuro psicólogo, este es un manual imprescindible por su calidad y amplitud. Aunque aparentemente no es un libro barato, podemos asegurar que es una inversión segura y altamente rentable. En cualquier caso, para los que tengan dudas, ofrecemos unos breves extractos del libro para que pueda convencerse por sí mismo. E. Barrull, 2001.
PrólogoTal como lo dan a entender las pinturas rupestres de la cubierta de este libro, la ecología es probablemente la ciencia más antigua, si no la profesión más antigua. Poco después de haber tenido sus primeros pensamientos conscientes, nuestros antepasados primitivos debieron darse cuenta de la importancia de saber los lugares en que podían encontrar plantas comestibles y animales para cazar, y en que podían encontrarse a salvo de los enemigos que pretendían atacarles. Pero hoy en día podemos llegar mucho más lejos. Sabemos que si deseamos utilizar los productos del mundo natural sin destruirlos, si deseamos producir alimento para nosotros y no para las plagas, y si deseamos predecir «qué sucederá después» en nuestro medio ambiente, hemos de comprender el mundo natural y los organismos que lo forman. Este libro ha sido pensado para documentar y fomentar dicha comprensión. Es un libro acerca de la distribución y la abundancia de los distintos tipos de organismos sobre la faz de la Tierra, y acerca de los caracteres e interacciones físicos, químicos y especialmente biológicos que determinan dichas distribuciones y abundancias. A diferencia de lo que sucede con otras ciencias, la materia de estudio de la ecología resulta evidente a todo el mundo; la mayoría de las personas han observado y ponderado la Naturaleza, y por ello se puede decir que todos somos ecólogos en cierta medida. Pero la ecología no es una ciencia fácil, ya que presenta grandes sutilezas y complejidades. Trabaja explícitamente en tres niveles de la jerarquía biológica —los organismos, las poblaciones de organismos y las comunidades de poblaciones— y, como veremos luego, ignora por su cuenta y riesgo los detalles de la biología de los individuos o las influencias de los acontecimientos históricos, evolutivos y geológicos. Utiliza los adelantos en los conocimientos de la bioquímica, el comportamiento, la climatología, la tectónica, etc., pero al mismo tiempo contribuye también al conocimiento de extensas áreas de la biología. Si, como dijo T. H. Dobzhansky, «en la biología nada tiene sentido si no es a la luz de la evolución», en la evolución muy pocas cosas tienen sentido si no es a la luz de la ecología; es decir, en términos de las interacciones entre los organismos y sus ambientes físicos, químicos y biológicos. La ecología presenta así mismo la característica de verse particularmente confrontada con la singularidad: millones de especies diferentes, incontables billones de individuos genéticamente distintos, todos ellos viven y se influyen mutuamente en un mundo variado y siempre cambiante. La belleza de la ecología estriba en que nos exige que desarrollemos nuestro conocimiento sobre problemas muy básicos y evidentes de un modo que reconozca el carácter único y la complejidad de todos los aspectos de la Naturaleza, pero que busque esquemas y predicciones dentro de esta complejidad en lugar de ser abrumado por ella. Tal como ha señalado L. C. Birch, la receta del doctor Whitehead para la ciencia no es nunca tan apropiada como al aplicarla a la ecología: busca la simplicidad, pero desconfía de ella. En este libro hemos enfocado el estudio de la ecología pasando de los organismos a las poblaciones, y de éstas a las comunidades. Pero en algunas ocasiones, y especialmente en los capítulos 14 y 15, hemos mirado hacia atrás para poder ver la luz que las interacciones más complejas arrojan sobre los niveles más simples de la jerarquía. La ecología no es una ciencia con una estructura lineal simple: todo afecta a todo lo demás. Los distintos capítulos contienen proporciones diferentes de historia natural descriptiva, fisiología, comportamiento, experimentación rigurosa de campo y de laboratorio, cuidadosos análisis y censos de campo, y modelos matemáticos (una forma de simplicidad que es esencial que se intente conseguir, pero de la que es igualmente esencial desconfiar). Estas proporciones variables reflejan, hasta cierto punto, los progresos efectuados en las diferentes áreas. Reflejan también diferencias intrínsecas de varios aspectos de la ecología. Por muchos progresos que se consigan, la ecología continuará siendo el lugar de encuentro del naturalista, del experimentador, del biólogo de campo y del especialista en modelos matemáticos. Creemos que todos los ecólogos deberían por lo menos intentar una combinación de todas estas facetas. El presente libro va destinado a todos aquellos cuyo programa de estudios incluye la ecología. Algunos aspectos del tema, en particular los matemáticos, resultarán difíciles para ciertos estudiantes, pero nuestro enfoque de la materia está pensado para asegurar una visión equilibrada y actual de la misma, sea cual fuere el punto fuerte del lector —el estudio de campo o de laboratorio, la teoría o la práctica. Una característica técnica del libro consiste en la incorporación de notas al margen, a modo de postes indicadores a lo largo de todo el texto. Creemos que con ello se consiguen varias finalidades. En primer lugar, estas notas constituyen una serie de subtítulos que subrayan la estructura detallada del texto. Sin embargo, puesto que son numerosas y a menudo contienen información por derecho propio, también pueden ser leídas una tras otra, junto con los subtítulos convencionales, para obtener un esquema de cada capítulo. Del mismo modo pueden servir también como ayuda para los estudiantes en el momento de repasar un tema —de hecho, son parecidas a las anotaciones que los propios estudiantes añaden a sus libros de texto—. Finalmente, puesto que las notas marginales suelen resumir el mensaje básico del párrafo o los párrafos que acompañan, pueden servir para la comprobación continuada de la comprensión: si el lector observa que la nota marginal corresponde al mensaje básico de lo que acaba de leer, puede deducir que lo ha comprendido. Una vez completado este libro, es un placer para nosotros manifestar nuestra gratitud a todas las personas que, de distintos modos, nos han ayudado a concebirlo, desarrollarlo y publicarlo. Numerosos colegas leyeron y comentaron diversos borradores del texto. John Lawton y Bob May se tomaron el trabajo de examinar todos los capítulos. Jo Anderson, Roy Anderson, David Atkinson, Leo Buss, Arthur Cain, James Chubb, Andrew Cossins, Robert Cowie, Mi-chael Crawley, John Parrar, Michael Hassell, William Heal, Alan Hildrew, Mi-chael Hutchings, Robert James, Richard Law, Richard Palmer, Mary Price, Derek Ranwell, Persio de Souza Santos Jr., Dave Thompson, Des Thompson, James Thomson, Jeff Waage, Richard Wall, Hilary Wallace, Andrew Watkinson y John Whittaker nos prestaron su ayuda para uno o varios capítulos. En la Blackwell Scientific Publications tuvimos la suerte de disponer de la ayuda y el estímulo de Simón Rallison, quien tomó nuestro texto y lo convirtió en un libro, y de Bob Campbell, quien nos halagó y animó durante todo el proceso. Andy Sinauer, de Sinauer Associates, nos proporcionó información sobre la reacción ante nuestro propósito de diversos ecólogos norteamericanos, y coordinó la revisión de los capítulos en Norteamérica. Nuestros apuntes se convirtieron en un texto gracias al trabajo paciente y cuidadoso de mecanografiado de Anita Callaghan, Jane Farrell, Gweneth Kell, Susan Scott y Maria Wooley. Finalmente, deseamos expresar nuestro agradecimiento a nuestras esposas y nuestras familias por habernos apoyado, escuchado e ignorado según lo requirieran las circunstancias.
Introducción: la ecología y su campo de acción La palabra «ecología» fue utilizada por primera vez por Ernest Haeckel en 1869. Parafraseando a Haeckel, podemos definir la Ecología como el estudio científico de las interacciones entre los organismos y su medio ambiente. La palabra deriva del griego oikos, que significa «hogar». Por consiguiente, se podría decir que la ecología es el estudio de la «vida doméstica» de los organismos vivos. Krebs (1972) ha sugerido una definición mucho menos vaga, más llena de información: «Ecología es el estudio científico de las interacciones que determinan la distribución y la abundancia de los organismos.» Esta definición posee el mérito de resaltar el tema fundamental de la ecología: la distribución y abundancia de los organismos —dónde se presentan, en qué número y qué hacen—. Sin embargo, la definición de Krebs no utiliza la palabra «ambiente», y para comprender la razón de ello es necesario definir esta palabra. El medio ambiente de un organismo consiste en todos aquellos factores y fenómenos externos al organismo y que influyen sobre él, ya se trate de factores físicos o químicos (abióticos) o bien de otros organismos (bióticos). Pero, naturalmente, las interacciones de que habla la definición de Krebs son interacciones con estos factores. Por consiguiente, el medio ambiente conserva también aquí la posición central que le concedió Haeckel en su definición de la ecología. En lo que se refiere al tema central de la ecología, la frase «la distribución y abundancia de los organismos» es agradablemente sucinta. Pero necesitamos ampliarla. La ecología tiene tres niveles de interés: el organismo individual, la población (formada por individuos de la misma especie) y la comunidad (formada por un número más o menos elevado de poblaciones). A nivel del organismo, la ecología se ocupa del modo en que los individuos se ven afectados por su ambiente biótico y abiótico (y por el modo en que influyen sobre éste). A nivel de la población, la ecología se ocupa de la presencia, o ausencia, de unas especies determinadas, de su abundancia o escasez, y de las oscilaciones y fluctuaciones de su número. Pero a este nivel existen dos enfoques. Uno de ellos se ocupa primero de las características de los organismos individuales, y luego estudia el modo en que estas características se combinan para determinar las de la población. El otro enfoque estudia directamente las características de las poblaciones, e intenta relacionarlas con los aspectos del medio ambiente. Ambos enfoques tienen su utilidad, y ambos serán utilizados. De hecho, estos dos enfoques son útiles también en el nivel de la comunidad. La ecología de las comunidades estudia la composición o estructura de las comunidades, y las vías seguidas por la energía, los nutrientes y otros productos químicos a medida que atraviesan dichas comunidades (el funcionamiento de las comunidades). Podemos intentar comprender estos esquemas y procesos partiendo del estudio de las poblaciones que componen la comunidad; pero también podemos estudiar directamente las propiedades de las comunidades, tales como la diversidad de especies, la tasa de producción de biomasa, etc. También en este caso se utilizarán los dos enfoques. La ecología es una disciplina central de la biología, y por ello no resulta sorprendente que muestre coincidencias con otras muchas disciplinas, especialmente con la genética, la evolución, el comportamiento y la fisiología. Sin embargo, el tema particular de la ecología se centra en aquellas características que afectan en mayor grado a la distribución y la abundancia —los procesos de natalidad, mortalidad y migración. Es importante señalar ya desde el
principio que los ecólogos estudian no sólo las comunidades, poblaciones y
organismos de la naturaleza, sino también los ambientes producidos por
el hombre o influidos por él (huertas, campos de trigo, graneros, reservas
naturales, etc.). Y más aún, los ecólogos se interesan a menudo por los
sistemas de laboratorio y los modelos matemáticos, que han desempeñado un
papel crucial en el desarrollo de la ecología y que están destinados a
continuar desempeñándolo. Serán examinados con frecuencia a lo largo del
presente libro. De todos modos, lo que nos interesa en último término es el
mundo real, y el valor de los modelos y de los simples experimentos de
laboratorio debe ser juzgado siempre en términos de la luz que arrojan sobre el
funcionamiento de los sistemas más naturales. Son un medio de llegar a un fin
—no son nunca un fin en sí mismos—. Uno de los objetos principales de la
ciencia estriba en simplificar, y con ello facilitar la comprensión de la
complejidad del mundo real. La ecología es una ciencia que apuntala a las
ciencias naturales e intenta explicarlas. Explicación, descripción, predicción y control En todos los niveles de la jerarquía ecológica podemos intentar que se efectúen varias cosas distintas. En primer lugar, podemos intentar explicar o comprender. Se trata de la búsqueda del conocimiento en la tradición científica pura. Mas para conseguirlo, primero es necesario efectuar una descripción. También con ello aumentamos nuestros conocimientos del mundo vivo. Es evidente que para comprender algo debemos disponer primero de la descripción de lo que deseamos comprender. Igualmente, aunque esto es menos obvio, las descripciones más valiosas son aquellas que se realizan teniendo en cuenta un problema particular o una «necesidad de comprender». Todas las descripciones son selectivas, pero es frecuente que en las descripciones no dirigidas, no realizadas con un fin determinado, resulte a posteriori que se han seleccionado unos factores equivocados. Los ecólogos tratan también con frecuencia de predecir lo que le sucederá a un organismo, a una población o a una comunidad bajo un conjunto determinado de circunstancias; y sobre la base de estas predicciones intentamos controlarlas o explotarlas. Intentamos minimizar los efectos de las plagas de langostas prediciendo cuándo es probable que se produzcan y aplicando las medidas apropiadas. Intentamos proteger las cosechas prediciendo en qué momento las condiciones serán favorables para la cosecha y desfavorables para sus enemigos. Intentamos preservar las especies raras prediciendo las medidas de conservación que nos permitirán conseguirlo. En algunos casos, la predicción y el control pueden ser efectuados sin disponer de explicaciones o comprensión. Pero las predicciones fiables, las predicciones exactas y las predicciones acerca de lo que sucederá en circunstancias no habituales sólo pueden ser conseguidas cuando podemos explicar lo que está sucediendo. Es importante subrayar que existen dos clases diferentes de explicación en la biología: explicaciones próximas y explicaciones remotas. Por ejemplo, la distribución y abundancia actuales de una determinada especie de ave puede ser «explicadas» en términos del ambiente físico que el ave tolera, del alimento que come y de los parásitos y depredadores que le atacan. Esta es una explicación próxima. Sin embargo, también podemos preguntarnos la razón de que esta especie de ave haya llegado a poseer las propiedades que actualmente parecen gobernar su vida. Esta pregunta debe ser contestada con una explicación en términos evolutivos. La explicación remota de la distribución y abundancia actuales de dicha ave se halla en las experiencias ecológicas de sus antepasados. En la ecología existen muchos problemas que requieren una explicación remota, evolutiva. «¿Cuál es la razón de que las especies coexistentes sean a menudo similares, pero rara vez idénticas?» (capítulo 7). «¿Por qué los depredadores adoptan unos esquemas particulares de comportamiento depredador?» (capítulo 9). «¿Cómo han llegado los organismos a poseer unas combinaciones de tamaño, tasa de desarrollo, nivel reproductor, etc.?» (capítulo 14). Estos problemas constituyen una parte de la ecología moderna, al igual que la prevención de las plagas, la protección de las cosechas y la conservación de las especies raras. Nuestra capacidad de control y explotación no puede más que mejorar con un incremento de nuestra capacidad de explicar y comprender. En la búsqueda del conocimiento debemos combinar las explicaciones próximas y remotas.
Introducción Hemos decidido empezar este libro con los capítulos dedicados a los organismos, para estudiar luego los modos en que se afectan mutuamente, y para considerar finalmente las propiedades de las comunidades formadas por dichos organismos. También habríamos podido enfocar el tema al revés — empezar con el estudio de las comunidades complejas de los hábitats naturales y creados por el hombre, analizarlas luego a escalas cada vez menores y terminar con los capítulos dedicados a las características de los distintos organismos. El problema se parece un poco al que se plantearía al escribir un libro sobre los relojes. Podríamos empezar con un estudio y una clasificación de los distintos tipos de relojes, y terminar con unos capítulos que hablaran sobre resortes, ruedas dentadas, pilas y cristales. Hemos elegido empezar con los organismos ya que sobre ellos actúan las fuerzas evolutivas (viven o mueren, dejan o no descendencia); y la naturaleza de las comunidades debe ser explicada en último término (más que simplemente descrita) en función de sus partes —poblaciones individuales compuestas por organismos individuales. Estudiamos primero los tipos de correspondencias que podemos detectar entre los organismos y los ambientes en que viven. Resultaría vulgar y superficial empezar con la idea de que cada organismo está amoldado idealmente, de algún modo, para vivir allí donde lo hace. Podemos llegar a esta conclusión tras un estudio exhaustivo, pero lógicamente no puede ser una premisa. De todos modos, subrayamos que los organismos son tal como son, y viven donde viven, a causa de su historia evolutiva (capítulo 1). Pasamos luego a estudiar el modo en que las condiciones ambientales varían de un lugar a otro y de un mohiento a otro, y las maneras en que los organismos se diferencian por cuanto a las condiciones que toleran o prefieren (capítulo 2). A continuación analizaremos los recursos que utilizan los diferentes tipos de organismos, y la naturaleza de su interacción con dichos recursos (capítulo 3). Los tipos de especies que se hallan presentes en una comunidad, sus números y/o sus masas respectivas, confieren a dicha comunidad una gran parte de su interés ecológico. La abundancia viene determinada por el equilibrio entre los nacimientos y las muertes, y entre la inmigración y la emigración. En el capítulo 4 estudiamos algunos de los diversos esquemas de nacimientos y muertes que, más adelante, comprobaremos que ejercen un profundo efecto sobre el comportamiento de las poblaciones. Una cierta capacidad de dispersión caracteriza a todas las especies vegetales y animales. Determina la tasa con que los organismos escapan de los ambientes que son desfavorables o que están empeorando, y la tasa de descubrimiento de los ambientes favorables para ser colonizados y explotados. La dispersión de los organismos se estudia en el capítulo 5.
El ajuste entre los organismos y su
ambiente 1.1 Introducción La ecología estudia los organismos y
su medio ambiente, y es importante que comprendamos la relación existente entre
ellos. La afirmación más fundamental que se puede hacer probablemente acerca
de dicha relación es que los distintos tipos de organismos no están
distribuidos al azar entre los diferentes tipos de ambientes: existe una
correspondencia entre ambos. Esta correspondencia forma parte de nuestro sentido
del orden de las cosas. Pero, ¿cuál es exactamente la naturaleza del ajuste
entre los organismos y sus ambientes respectivos? 1.1.1 Selección natural — ¿adaptación o abaptación? En cualquier conferencia, la frase que se emplea más a menudo para describir este ajuste es: «el organismo X está adaptado a», seguida de una descripción de las condiciones en las que se encuentra a dicho organismo. Así, podemos oír que «los peces están adaptados a la vida en el agua», o que «los cactus están adaptados a las condiciones de sequía que reinan en los desiertos», etc. Paradójicamente, el modo más seguro de comprender la verdadera naturaleza del ajuste entre los organismos y sus ambientes consiste en aclarar la razón de que «adaptado a» es una expresión totalmente inapropiada en este contexto.
La teoría de Charles Darwin (1859) sobre la evolución por selección natural puede ser considerada, simplificadamente, como basada en una serie de proposiciones. I. Los individuos que constituyen una población de una especie no son idénticos: muestran variaciones, a veces muy reducidas, en cuanto a tamaño, tasa de desarrollo, respuesta a la temperatura, etc. II. Por lo menos una parte de dicha variación es hereditaria. En otras palabras, las características de un individuo están determinadas hasta cierto punto por su dotación genética. Los descendientes reciben sus genes de sus progenitores, y por consiguiente la descendencia muestra una tendencia a compartir ciertas características con los progenitores. III. Todas las poblaciones poseen la potencialidad de poblar toda la Tierra, y lo harían si todos los individuos sobrevivieran y si cada individuo produjera el número máximo de descendientes. Pero no lo hacen: muchos individuos mueren antes de reproducirse, y la mayoría de ellos (si no todos) se reproducen con una tasa inferior a la máxima. IV. Los distintos individuos dejan un número distinto de descendientes. Esto significa más que decir que los diferentes individuos tienen distinto número de descendientes. Incluye también las posibilidades de supervivencia V. Finalmente, el número de descendientes que deja un individuo depende, no por completo, pero sí de modo crucial, de la interacción entre las características del individuo y el medio ambiente del mismo. Un individuo sobrevivirá, se reproducirá y dejará descendientes en ciertos ambientes, pero no en otros. En este sentido se podría decir que la Naturaleza es selectiva. En este sentido algunos ambientes pueden ser descritos como favorables o desfavorables, y en este mismo sentido algunos organismos pueden ser considerados aptos o no. Si, a consecuencia de que algunos. individuos dejen más descendencia que otros, las características hereditarias, de una población cambian de generación en generación, se dice que se ha producido una evolución por selección natural. Sería razonable decir que los
organismos de una determinada generación están abaptados por los
ambientes de las generaciones anteriores. Los medios ambientes anteriores actúan
a modo de filtro, a través del que han pasado las combinaciones de caracteres
hasta llegar al momento actual. Pero parece que los organismos están adaptados
(ajustados) a su medio ambiente actual únicamente porque dicho medio ambiente
tiende a ser similar al ambiente anterior. La palabra «adaptación» provoca
una impresión errónea de predicción, de previsión o, en último término, de
intención. Los organismos no están previstos para el presente o el futuro, ni
están adaptados a ellos —son consecuencias del pasado, y por consiguiente han
sido adaptados por éste. 1.1.2 Eficacia Por definición, los individuos más aptos de una población son aquellos que dejan un mayor número de descendientes. En la práctica, el término es aplicado con frecuencia no a un solo individuo, sino a un individuo típico o a un tipo: por ejemplo, se puede decir que en las dunas de arena, los caracoles de concha amarilla son más aptos que los de concha parda (es decir, tienen más probabilidades de sobrevivir y de dejar un mayor número de descendientes).
Pero la aptitud o eficacia es un término relativo, no absoluto. El número de semillas producido por una planta, o el de huevos producido por un insecto, no es una medida directa de su eficacia; ni tampoco lo es el número de descendientes que dejan. Lo que determina su eficacia es más bien la contribución proporcional de un individuo a las generaciones futuras: los individuos más aptos de una población son los que dejan un mayor número de descendientes en relación con el número de descendientes de otros individuos, menos eficaces, de la población. Y aquellos individuos que dejan la mayor proporción de descendientes en una población ejercen una mayor influencia sobre las características hereditarias de dicha población.
Ninguna población de organismos puede contener todas las posibles variantes genéticas que podrían existir e influir sobre la eficacia. De ello se deduce que es poco probable que la selección natural conduzca a la evolución de individuos perfectos, de «aptitud máxima». Favorece a los más aptos de entre los individuos existentes, y éste puede ser un campo muy restringido. Los poderes de las fuerzas evolutivas no son ilimitados, ni mucho menos. La teoría de Darwin no predice la perfección, ni tan siquiera en un medio ambiente que no experimente cambios de generación en generación. Tan sólo predice que algunos individuos dejarán más descendientes que otros, y por ello ejercerán una mayor influencia en la determinación de las características de las generaciones futuras. Esta es la limitación fundamental al grado en que los organismos se ajustan a su medio ambiente. La esencia misma de la selección natural estriba en que los organismos llegan a ajustarse a sus ambientes siendo «los más aptos existentes» o «los más aptos por el momento»: no son «los mejores que se podría imaginar». Las razones por las que no esperamos encontrar la evolución de la perfección en la Naturaleza se hallan estudiadas más a fondo en los trabajos de Jacob (1977), Gould & Lewontin (1979) y Harper (1982). Existen además otras limitaciones y condiciones en cuanto al modo y al grado en que los organismos se ajustan a sus ambientes. Algunas de ellas serán estudiadas en este capítulo. 1.2 Factores históricos Es particularmente importante tener en
cuenta que los acontecimientos pasados, ocurridos en la Tierra, pueden tener
profundas repercusiones para el presente. Nuestro mundo no ha sido construido
por alguien que tomó uno a uno a los distintos organismos y los examinó con
respecto a cada uno de los ambientes con el fin de que cada organismo encontrara
su lugar perfecto. Es un mundo en el que los organismos viven donde lo hacen por
razones que son a menudo, por lo menos en parte, accidentes de la historia. La curiosa distribución de los organismos entre los continentes, aparentemente inexplicable en términos de dispersión a gran distancia, condujo a los biólogos, y especialmente a Wegener (1915), a sugerir que debieron ser los propios continentes los que se desplazaron. Esta idea fue rebatida por los geólogos hasta que las mediciones geomagnéticas exigieron la misma explicación, en apariencia altamente improbable. El movimiento de las placas tectónicas de la corteza terrestre, con la consiguiente migración de los continentes, reconcilia a los geólogos y biólogos (fig. 1.1). Así, mientras los reinos vegetal y animal estaban sufriendo grandes desarrollos evolutivos, las poblaciones fueron divididas y separadas, y las áreas de tierra se desplazaron a través de las zonas climáticas.
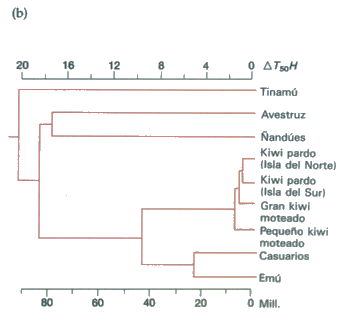
La figura 1.2 muestra un ejemplo de un grupo importante de organismos (las grandes aves no voladoras) cuyas distribuciones sólo empiezan a tener sentido a la luz del movimiento de las masas de tierra. Sería injustificado decir que los emúes y los casuarios se hallan allí donde están porque representan la mejor adaptación a los ambientes australianos, mientras que el ñandú y el tinamú se encuentran allí donde viven porque a su vez representan la mejor adaptación a los ambientes sudamericanos. Cada una de estas aves ha evolucionado en su propio continente y ha sido abaptada por los ambientes existentes allí en el pasado. Pero sus distribuciones muy dispares están determinadas esencialmente por los movimientos prehistóricos de los continentes y por la subsiguiente imposibilidad para cada una de ellas de llegar al medio ambiente de las otras aves.
Figura 1.2 (a) La distribución y el grado de afinidad de un grupo de grandes aves no voladoras pueden ser explicados, por lo menos en parte, por la deriva continental (véase la figura 1.1). (b) Los grados de afinidad pueden ser medidos con una técnica de hibridación del DNA. El DNA de doble hebra es dividido en dos hebras separadas mediante la aplicación de calor. A continuación se pueden combinar las hebras de especies diferentes, separándolas luego de nuevo por calor. Cuanto más similares son las dos hebras, tanto más elevada será la temperatura necesaria para separarlas, DTT50H. La temperatura de separación proporciona una medida del parentesco de las especies y un valor estimativo del momento en que divergieron. Mili = millones de años. La primera divergencia fue la de los tinamúes con respecto a las restantes especies (agrupadas bajo el nombre de Ratitae). Las divergencias subsiguientes concuerdan con las fases de la rotura de Gondwanaland y la subsiguiente deriva continental (figura 1.1): (i) la grieta entre Australia y los demás continentes meridionales; (ii) la aparición del Atlántico entre África y Sudamérica; (iii) la aparición del Mar de Tasmania hace unos 80 mili, seguida probablemente por una migración a través de las islas de los antepasados del kiwi hasta Nueva Zelanda hace 40 mili. La divergencia de las diversas especies de kiwis parece ser muy reciente. (Según Diamond, 1983, a partir de datos de Sibley y Ahlquist.) Continúa ...
Introducción La actividad de todo organismo cambia el medio ambiente en el que vive. Puede alterar las condiciones — como sucede cuando el movimiento de una lombriz de tierra a través del suelo causa la ventilación del mismo, o cuando la transpiración de un árbol refresca la atmósfera. O puede añadir o sustraer recursos del ambiente y alterar así su disponibilidad para otros organismos — como sucede cuando un árbol arroja sombra sobre lo que está debajo de él, o cuando una vaca come la hierba de un prado. Pero además, los organismos entran realmente en interacción unos con otros cuando los individuos, de un modo u otro, penetran en la vida de los demás. En los capítulos siguientes (6-13) estudiaremos estas interacciones. Distinguiremos cinco categorías principales: competencia, depredación, parasitismo, mutualismo y detritovorismo. Al igual que la mayoría de las categorías biológicas, tampoco éstas son perfectamente claras. En términos muy amplios, la «competencia» es una interacción en la que un organismo consume un recurso que habría estado disponible para otro organismo y habría podido ser consumido por éste. Un organismo priva a otro de algo y, como consecuencia, este otro organismo crece más lentamente, deja menos descendientes o corre un mayor peligro de morir. El acto de desposeimiento puede ocurrir entre dos miembros de la misma especie o entre individuos de diferentes especies. Hemos decidido estudiar estas situaciones en capítulos distintos (capítulos 6 y 7), en parte porque parece razonable suponer que los miembros de la misma especie tienen muchas probabilidades de buscar la satisfacción de las mismas necesidades en el ambiente (y de reaccionar simétricamente a la presencia uno del otro), mientras que los miembros de diferentes especies es probable que se diferencien en este aspecto (y que reaccionen asimétricamente a la presencia uno de otro). Pero es importante no tomar demasiado al pie de la letra esta distinción — el proceso de la selección natural depende de las diferencias entre miembros de la misma especie, y los individuos de una misma especie se pueden diferenciar también en lo referente a su estado de salud, su fase de desarrollo, etc. Además, los miembros de dos especies muy distintas pueden utilizar y agotar los mismos recursos: un conejo de un campo puede ser privado de su alimento con más eficacia por una oveja que por otro conejo. Los capítulos 8, 9 y 10 están dedicados a los diversos aspectos de la «depredación», aunque hemos tomado una definición muy amplia de depredación. Hemos combinado aquellas situaciones en las que un organismo come a otro y lo mata (por ejemplo, una lechuza capturando a un ratón), y aquellas en las que el consumidor toma sólo una parte de su presa, que puede crecer de nuevo para suministrar más tarde otro bocado (ramoneo). También hemos combinado el herbivorismo (los animales se comen las plantas) y el carnivorismo (los animales comen a otros animales). En el capítulo 8 examinamos la naturaleza de la depredación, es decir lo que le sucede al depredador y a la presa (ya sea ésta animal o vegetal). Se presta una especial al herbivorismo debido a las sutilezas que caracterizan la respuesta de una planta a los ataques. En el capítulo 9 estudiamos el comportamiento de los depredadores —campo en que se fusionan la ecología demográfica y la ecología del comportamiento. El modo en que se comportan los depredadores individuales influye claramente sobre la dinámica de las poblaciones del depredador y de la presa, y estas «consecuencias del consumo» constituyen la materia del capítulo 10. Esta es la parte de la ecología que tiene mayor importancia para aquellas personas dedicadas a la administración de los recursos naturales — la eficacia de la explotación (ya sea de peces, ballenas, pastos o prados) y el control biológico y químico de las plagas y malas hierbas. Cuatro de los procesos estudiados en esta sección —competencia, depredación, parasitismo y mutualismo— implican la presencia de unas interacciones entre organismos que generan procesos de selección natural. Cabe esperar que las actividades de los depredadores influyan sobre la evolución de sus presas, y viceversa; y cabe esperar también que las actividades de los parásitos influyan sobre la evolución de sus huéspedes, y viceversa. Si el proceso de la evolución es en gran parte consecuencia de las interacciones entre los organismos (tal como suponía Darwin), es entre los depredadores y sus presas, los parásitos y sus huéspedes, las parejas de mutualistas y entre los competidores donde podemos esperar que los procesos ecológicos y evolutivos se nos presenten más íntimamente relacionados. En cambio, la materia viva no deja descendientes y no puede evolucionar, ni tampoco multiplicarse. De todos modos, se halla implicada en interacciones complejas, y los procesos por los que es descompuesta incluyen competencia, parasitismo, depredación y mutualismo: microcosmos de todos los principales procesos ecológicos (salvo la fotosíntesis). El detritivorismo, el consumo de la materia orgánica muerte, se estudia en el capítulo 11. El capítulo 12, titulado Parasitismo y enfermedad, se ocupa de un tema que rara vez merece la atención de un capítulo entero en los textos de ecología. Y sin embargo, más de la mitad de las especies de la Tierra son parásitas. La categoría del parasitismo tiene unos límites poco estrictos, particularmente allí donde se fusiona con la depredación. Pero mientras un depredador suele tomar una parte o todo el cuerpo de muchas presas individuales, un parásito suele obtener sus recursos de uno o muy pocos huéspedes y (al igual que los depredadores remoneadores), rara vez mata a su huésped inmediatamente, si es que llega a matarlo. Los insectos parasitoides [como los icneumónidos que se desarrollan (a menudo en número de uno solo) en el cuerpo del huésped y lo matan] no encajan ni entre los depredadores ni entre los parásitos. Hablaremos de ellos, junto con los depredadores y los ramoneadores, en los capítulos 8-10. Finalmente, el capítulo 13 está dedicado a las interacciones mutualistas entre individuos de distintas especies. Se trata de interacciones de las que ambos organismos obtienen un beneficio neto. La ecología del mutualismo ha sido descuidada y, al igual que el parasitismo, rara vez consigue un capítulo entero en un libro de texto de ecología. Pero la mayor parte de la biomasa del mundo está compuesta por mutualistas. Las interacciones entre los organismos han sido a menudo resumidas mediante un código simple que representa a cada uno de los dos organismos de la interacción con los signos «+», «—» o «0», según como se vea afectado por la interacción. Así, una interacción depredador-presa (incluida una interacción herbívoro-presa), en la que el depredador se beneficia y la presa es perjudicada, se simboliza con + —; y la interacción parásito-huésped es también claramente + —. Otro caso claro es el del mutualismo, que es + +; si los organismos no muestran ningún tipo de interacción, podremos escribir 00 (situación denominada a veces «neutralismo»). El detritivorismo debe ser simbolizado por +0, ya que el detritívoro sale beneficiado, mientras que su alimento (ya muerto) no queda afectado por la relación. El término general aplicado a las interacciones +0 es el de «comensalismo», pero paradójicamente este término no suele ser utilizado para los detritívoros. En vez de ello es reservado para aquellos casos, afines al parasitismo, en que un organismo («el huésped») suministra recursos o cobijo a otro organismo, pero sin sufrir un perjuicio tangible. La competencia es a menudo descrita como una interacción — —, pero como veremos en los capítulos 6 y 7, a menudo resulta imposible establecer que ambos organismos salen perjudicados. Estas interacciones asimétricas pueden aproximarse entonces a una descripción —0, a la que se suele dar el nombre de «amensalismo». Los verdaderos casos de amensalismo se pueden producir cuando un organismo provoca su efecto perjudicial (por ejemplo, una toxina) tanto si el organismo potencialmente afectado se halla presente como si no. Los ocho capítulos de esta sección tienen estilos bastante distintos, que son un reflejo de las diferencias existentes en el «estado de la materia». Los estudios sobre el parasitismo y el mutualismo se han centrado sobre todo en desentrañar los misterios de las relaciones entre los organismos en cuestión, en lugar de considerar el papel de los fenómenos en las comunidades en que viven los organismos. Así, sabemos mucho acerca de la ecología de las bacterias fijadoras de nitrógeno que viven en los nódulos de las raíces de las leguminosas, pero sabemos mucho menos sobre la ecología de las leguminosas con nódulos en la vegetación natural. Análogamente, conocemos a fondo la ecología de los gusanos parásitos del intestino, pero sabemos muy poco acerca del papel que desempeña la infección en la determinación de la abundancia de las poblaciones naturales. Por otro lado, en el caso de la competencia interespecífica, el problema es algo distinto. Tenemos una ciencia muy sofisticada dedicada al estudio experimental y teórico de la competencia, pero ignoramos aún en gran medida la frecuencia con que se produce la competencia y la importancia que tiene como fuerza en las comunidades naturales. También existe un importante contraste entre la larga historia de modelos matemáticos de las interacciones depredador-presa y la reciente atención que se está presentando a los modelos del mutualismo. Por razones de este tipo, y también debido a los intereses especiales de los investigadores más influyentes, el estudio ecológico progresa a tontas y a locas. Existen partes de la ecología que disponen de una gran base teórica, pero carecen del apoyo de la historia natural, mientras que otras partes consisten sobre todo en anécdotas de la historia natural y no disponen de una buena teoría que les proporcione coherencia. La ecología es una ciencia que está llena de lagunas y de áreas de ignorancia —al igual que cualquier otra ciencia.
6. Competencia
intraespecífica
Los organismos crecen, se reproducen, mueren y emigran (capítulos 4 y 5). Se ven afectados por las condiciones en que viven (capítulo 2) y por los recursos que obtienen (capítulo 3). Pero ningún organismo vive aislado. Todos ellos, por lo menos durante una parte de su vida, son miembros de una población compuesta por individuos de su propia especie.
Los individuos de una misma especie tienen necesidades muy similares para sobrevivir, crear y reproducirse; pero la necesidad combinada de todos ellos por un recurso puede exceder la oferta del mismo. Los individuos compiten entonces por dicho recurso y por lo menos algunos de ellos quedan privados de él. El presente capítulo estudia la naturaleza de esta competencia intraespecífica, y sus efectos sobre los individuos competidores. Resultará útil empezar con una definición de trabajo de la competición: «competición es una interacción entre individuos, provocada por la necesidad común de un recurso limitado, y conducente a la reducción de la supervivencia, el crecimiento y/o la reproducción de los individuos competidores». Ahora podemos pasar a estudiar más a fondo la cuestión.
Consideremos primero una comunidad hipotética simple: una población próspera de saltamontes (todos de la misma especie) que vive sobre un campo de gramíneas (también de una sola especie). Para poder vivir, los saltamontes deben consumir hierba para conseguir energía y materia con que construir su cuerpo. Pero en el proceso de encontrar y consumir el alimento, utilizan también energía y se exponen al peligro que suponen los depredadores. Cada saltamontes se encontrará frecuentemente en un lugar del campo en que había existido previamente una brizna de hierba —es decir, antes de que otro saltamontes la comiera. Siempre que sucede esto, el saltamontes debe continuar buscando; debe gastar más energía y correr un riesgo mayor antes de poder comer. Y esto ocurrirá con mayor frecuencia cuantos más saltamontes se encuentren en el campo, compitiendo por la comida. Pero un aumento del gasto de energía, un mayor riesgo de mortalidad y una menor tasa de ingestión de alimento reducirán las probabilidades de supervivencia del saltamontes; además, el mayor gasto de energía y la menor ingestión de alimento pueden hacer que el individuo tenga menos energía disponible para su desarrollo y para la reproducción. Así, puesto que la supervivencia y la reproducción determinan la contribución de un saltamontes a la siguiente generación, cuantos más competidores intraespecíficos por el alimento tenga un saltamontes, tanto menos será dicha contribución por su parte. En lo que se refiere a la hierba, la
contribución de un genete individual a la generación siguiente dependerá del
número de sus descendientes que a su vez se desarrollen hasta adultos
reproductores. Una plántula aislada que crece en un suelo fértil puede tener
una probabilidad muy alta de sobrevivir hasta la madurez reproductiva.
Probablemente mostrará una gran cantidad de crecimiento modural, y por ello
producirá un gran número de descendientes. Sin embargo, una plántula que esté
estrechamente rodeada por plántulas vecinas (que le hacen sombra con sus hojas
y que agotan el suelo con sus raíces), tiene muy pocas probabilidades de
sobrevivir, y si lo consigue será seguramente pequeña y simple, y producirá
pocas semillas. Por consiguiente, el aumento de la densidad reducirá la
contribución que cada individuo hace a la generación siguiente. 6.2 Rasgos comunes de la competencia intraespecífica
Es evidente que estos dos casos de competencia intraespecífica muestran una serie de rasgos en común. El primero de ellos es que el efecto último de la competencia es una menor contribución a la generación siguiente, en comparación con lo que habría sido dicha contribución si no hubieran existido competidores. La competencia intraespecífica conduce a una disminución de las tasas de ingestión de recursos por individuo, quizás a una disminución de las tasas de crecimiento o desarrollo individual, o a una reducción de las reservas almacenadas. Esto puede conducir a su vez a una reducción de la supervivencia y/o de la fecundidad. Como vimos en el capítulo 4, la supervivencia y la fecundidad determinan en conjunto el output reproductor de un individuo.
El segundo rasgo de la competencia intraespecífica es que el recurso por el que compiten los individuos debe hallarse en cantidad limitada. El oxígeno, por ejemplo, si bien es un recurso absolutamente esencial, no es algo por lo que compitan los saltamontes ni las hierbas; la cantidad disponible supera la tasa con que pueden consumirlo las poblaciones más densas. Análogamente, la luz, el alimento, el espacio o cualquier otro recurso sólo son disputados por los competidores si se encuentran en cantidad limitada.
En muchos casos, los individuos competidores no interaccionan unos con otros directamente. Los individuos responden al nivel de un recurso, que ha sido disminuido por la presencia y la actividad de otros individuos. Así, los saltamontes que compiten por el alimento no se ven directamente afectados por otros saltamontes, sino por la reducción en el nivel de comida y por la creciente dificultad de encontrar buena comida que no haya sido ya explotada por los otros. Análogamente, una planta herbácea competitiva se ve afectada adversamente por la presencia de plantas próximas, ya que la zona de la que extrae los recursos (luz, agua, nutrientes) ha quedado afectada por las «zonas de privación de recurso» de las plantas vecinas. En todos estos casos, la competencia puede ser descrita como explotación, por cuanto cada individuo se encuentra afectado por la cantidad de recurso que queda después de haber sido explotado por los otros. Pero en otros muchos casos la competencia toma otra forma, conocida como interferencia. Los individuos interaccionan aquí directamente unos con otros, y un individuo impedirá realmente a otro que ocupe una porción del habitat y por lo tanto que explote los recursos que se hallan en él. Este caso se encuentra, por ejemplo, entre los animales móviles que defienden un territorio (estudiado con mayor detalle en la sección 6.11): el resultado es a menudo que el propio territorio se convierte en recurso. La interferencia se puede producir también entre organismos sésiles. La presencia de un balano sobre una roca, por ejemplo, impide que otro balano ocupe esta misma posición, si bien el alimento en este lugar puede exceder a la demanda de un sólo organismo. De hecho, la interferencia es muy frecuente entre los animales y plantas sésiles que viven en las costas rocosas: con frecuencia compiten a través del crecimiento de un individuo por encima de otro. En estos casos, los efectos de la competencia suelen ser obvios — en numerosos casos de explotación, los efectos son mucho más útiles.. En la práctica, la interferencia está casi siempre acompañada por un elemento de explotación aunque, evidentemente, existen numerosos casos de explotación sin interferencia.
El tercer rasgo de la competencia intraespecífica estriba en que los individuos competidores son en esencia equivalentes —pero no lo son en la práctica. El mismo hecho de que hayan sido clasificados como de «la misma especie» implica que poseen muchas características fundamentales en común, y que cabe esperar que utilicen recursos similares y reaccionen de modo muy parecido a las condiciones. Sin embargo, es importante no llevar demasiado lejos la idea de que los efectos entre los individuos competidores son recíprocos. Existen muchas ocasiones en las que la competencia intraespecífica es muy desequilibrada: una plántula precoz, robusta, probablemente hará sombra a otra plántula más tardía, enana, y un briozoo de más edad y mayor tamaño probablemente «crecerá por encima» de otro individuo más pequeño y joven. Además, las diferencias hereditarias entre los individuos pueden asegurar con certeza que las interacciones competitivas no son recíprocas. Los genotipos altos del maíz, por ejemplo, habitualmente harán sombra y suprimirán a los genotipos bajos de la misma especie. Por consiguiente, no podemos decir que los individuos competidores de una misma especie son enteramente equivalentes. Lo que podemos decir es que los miembros de la misma especie tienen más probabilidades que los miembros, de especies distintas de necesitar el mismo recurso y de reaccionar recíprocamente uno en la presencia del otro.
Esta falta de equivalencia exacta significa que el efecto final de la competición no es ni mucho menos igual en los distintos individuos. Los competidores débiles contribuirán poco a la generación siguiente, o no contribuirán en absoluto. Es posible que la contribución de los competidores fuertes se vea sólo muy poco afectada. De hecho, un competidor fuerte puede realizar incluso una contribución proporcional superior cuando existe una intensa competencia, en comparación con el caso en que no exista competencia (es decir, si mantiene su contribución cuando todos los individuos que le rodean disminuyen la suya). En otras palabras, aunque el efecto final de la competencia es una reducción del output reproductivo, ello no significa siempre una disminución de la eficacia (es decir, de la contribución relativa) individual, en especial en el caso de los competidores más fuertes. Por consiguiente, no sería correcto afirmar que la competencia «afecta adversamente» a todos los individuos competidores (Wall & Begon, 1985).
Finalmente, el cuarto rasgo de la competencia intraespecífica es que su efecto probable sobre cualquier individuo es mayor cuanto más elevado es el número de competidores. Por ello se dice que los efectos de la competencia intraespecífica dependen de la densidad. Para estudiar más a fondo la competencia intraespecífica debemos examinar los efectos de la densidad demográfica sobre los individuos, y en especial sus efectos sobre la muerte, el nacimiento y el crecimiento. Continúa ...
12. Parasitismo y enfermedad
En el capítulo 8 definíamos a un parásito diciendo que es un organismo que obtiene sus nutrientes de uno o unos pocos individuos huéspedes, provocándoles habitualmente un daño, pero sin causarles una muerte inmediata. Sin embargo, existen muchas definiciones de parasitismo. Los parasitólogos tienden a subrayar a) la intimidad de la asociación entre el parásito y el huésped, y b) la dependencia del parásito con respecto al huésped en cuanto a la regulación de su ambiente; pero pueden omitir cualquier mención al «daño» producido al huésped. Para el ecólogo es importante destacar esta intimidad de la asociación en una definición completa del parasitismo, pero si no existe beneficio ni daño real para el huésped, la asociación debe ser considerada como comensalismo (pág. 203). Por esta razón parece razonable distinguir entre interacciones de parasitismo, en las que se produce predeciblemente algún daño, aunque quizá tan sólo en circunstancias apropiadas (por ejemplo, número suficiente de parásitos o mal estado del huésped), e interacciones de comensalismo, en las que el organismo comensal sale beneficiado, pero sin que el huésped resulte beneficiado ni dañado. También necesitamos una definición operativa de «daño» o «perjuicio», si queremos que la definición de parasitismo nos resulte útil. En la práctica, podemos medir el perjuicio que un parásito ocasiona a su huésped como la reducción de la tasa intrínseca de crecimiento del huésped y/o de su población.
Los parásitos y patógenos (agentes causantes de enfermedades) son un grupo extraordinariamente importante de organismos. Millones de personas mueren cada año a causa de diversos tipos de infección, y muchos millones más quedan debilitadas o deformes (250 millones de casos de elefantiasis en la actualidad, más de 200 millones de casos de bilharziosis, y la lista no termina aquí). Si a ello se añaden los efectos de los patógenos sobre los animales domésticos y los cultivos, el coste en términos de sufrimiento humano y pérdidas económicas resulta incalculable. Naturalmente, los seres humanos les ponen las cosas fáciles a los parásitos y patógenos (véase más adelante) al vivir en poblaciones densas y agregadas y al obligar a los animales domésticos y a las plantas cultivadas a hacer lo mismo. Pero, de todos modos, es razonable creer que los animales y plantas en general son dañados y matados en gran número por el parasitismo y la enfermedad.
Es seguro que los patógenos representan una importante fuente de mortalidad y de fecundidad no realizada en muchas poblaciones naturales, probablemente en la mayoría de ellas.
Finalmente, en términos de número de individuos y de especies, basta con tener en cuenta dos cosas. En primer lugar, un organismo de vida libre que no albergue varios individuos parásitos de varias especies es un caso raro; y en segundo lugar, muchos parásitos y patógenos muestran especificidad de huésped o por lo menos poseen una gama limitada de huéspedes. En conjunto, estos dos puntos parecen hacer inevitable la conclusión de que más de la mitad de las especies de la Tierra, y mucho más de la mitad de los individuos, son parásitos o patógenos. Lo más probable es que la mayoría de los parásitos bacterianos y víricos aún no hayan sido identificados. A la vista de esta riqueza, puede
parecer irrealizable la tarea de efectuar un breve estudio de la diversidad de
parásitos y patógenos. Pero por otro lado, un resumen superficial,
especialmente de los tipos que han sido estudiados más a fondo y que tienen más
importancia para el hombre, constituirá una base de gran valor para el resto
del presente capítulo. 12.2 La diversidad de los parásitos El lenguaje y los términos especializados que emplean los fitopatólogos y los zooparasitólogos son a menudo tan diferentes que dan la impresión de que los temas tratados no tienen nada en común. Pero esta distinción resulta superficial para el ecólogo. Existen importantes diferencias en el modo en que los animales y plantas vivos sirven de hábitat para los parásitos, y también importantes diferencias en cuanto a sus respuestas al ataque de los parásitos; pero las diferencias son menos notables que las similitudes. Por consiguiente, como ecólogos, estudiaremos ambos temas a la vez. Es conveniente reconocer desde el principio dos categorías principales entre los parásitos: microparásitos y macroparásitos. La distinción entre micro y macroparásitos es relativamente reciente (May & Anderson, 1979). Los microparásitos se multiplican directamente dentro de sus huéspedes (habitualmente dentro de las células huésped). Los macroparásitos crecen en el interior de sus huéspedes, pero se multiplican produciendo fases infectivas que salen fuera del huésped para infectar a otros huéspedes. Son a menudo intercelulares (en las plantas) o viven en las cavidades corporales, más que en el interior de las células del huésped. Continúa ...
13.1 Introducción Mutualismo es el nombre que se da a las asociaciones entre pares de especies, con beneficios mutuos (+,+); los individuos de una población de cada especie mutualista crecen y/o sobreviven y/o se reproducen con una tasa superior cuando se hallan en presencia de individuos de la otra especie. Cada mutualista consigue alguna de una serie de ventajas diferentes. En la mayoría de los casos, esta ventaja reside en un recurso alimenticio para una de las dos partes, por lo menos, y con frecuencia reside, para la otra especie, en la protección contra los enemigos o la existencia de un ambiente favorable en el que crecer y reproducirse. En otros casos, la especie que obtiene alimento proporciona un «servicio» librando a su socio de sus parásitos (por ejemplo, el pez limpiador) o bien produciendo la polinización o la dispersión de las semillas. A pesar de las ventajas que obtiene cada socio, es importante no considerar al mutualismo como una relación «afectuosa» entre los mutualistas. Cada uno de ellos actúa de modo esencialmente «egoísta»; las relaciones mutualistas aparecen simplemente porque los beneficios que aportan a cada socio superan los costes que pudieran ocasionar. El lector encontrará un estudio detallado del tema en Boucher et al. (1984).
Los libros de ecología solían quitar importancia o ignorar por completo el mutualismo. Y sin embargo se trata de un fenómeno extraordinariamente difundido. Una parte muy significativa de la biomasa mundial depende de él —por ejemplo, los organismos dominantes de todos los prados y bosques (raíces de las plantas íntimamente asociadas con hongos en las micorrizas), y los corales (pólipos animales que contienen algas unicelulares) son mutualistas. La mayoría de los vegetales con raíces poseen micorrizas mutualistas; muchas plantas con flor dependen de los insectos polinizadores; y un gran número de animales contienen en sus intestinos una comunidad mutualista de microorganismos. El mutualismo ha sido muy pocas veces el objeto de los modelos matemáticos que han caracterizado el estudio de las interacciones competitivas, de las interacciones depredador-presa y parásito-huésped (May, 1982), aunque se están produciendo importantes cambios en este campo (Vandermeer & Boucher, 1978; Vandermeer, 1980). El estudio de la ecología de los mutualistas ha estado dominado por los temas relacionados con la biología general de los mutualismos (fisiología, morfología y comportamiento) más que con su dinámica de poblaciones. La estructura del presente capítulo refleja esta inclinación.
En primer lugar se consideran algunos ejemplos de lazos mutualistas entre individuos de diferentes especies que van desde aquellos que son facultativos [relaciones en las que cada individuo (simbionte) obtiene un beneficio, pero no depende del control del otro], pasando por los lazos que son obligados para uno de los individuos pero facultativos para el otro, hasta aquellos lazos que son obligados para ambos individuos. Luego estudiamos aquellos mutualismos en los que una especie vive en la superficie o en una cavidad corporal de la otra, así como las situaciones en las que un socio (denominado entonces endobionte) vive en el interior de las células del otro y en las que ambos componentes empiezan a mostrar una fisiología integrada (fig. 13.1). Hacia el final del capítulo discutiremos la idea según la cual ha sido la evolución de las relaciones mutualistas entre los organismos la que ha sido responsable de los principales pasos de la evolución de los animales y plantas superiores. También se intentará comparar la ecología de los parásitos con la de los mutualistas.
13.2 Mutualismos que implican vínculos recíprocos del comportamiento No resulta fácil catalogar los
beneficios obtenidos por los participantes de los diversos mutualismos que
implican intrincados vínculos de comportamiento. Su diversidad queda ilustrada
en los siguientes ejemplos. 13.2.1 El pájaro de la miel y el ratel Un pájaro africano, el pájaro guía
de la miel, ha establecido una notable relación con el ratel o tejón melífero.
Un pájaro de la miel que ha localizado un nido de abejas guía a un ratel hasta
el lugar. El- mamífero rompe el nido y se alimenta de la miel y de las larvas
de las abejas, y luego el pájaro obtiene su comida de cera y larvas. El pájaro
de la miel puede localizar un nido de abejas, pero no puede romperlo, mientras
que el ratel se encuentra en la situación inversa. El vínculo recíproco de
sus comportamientos les proporciona beneficios a ambos.
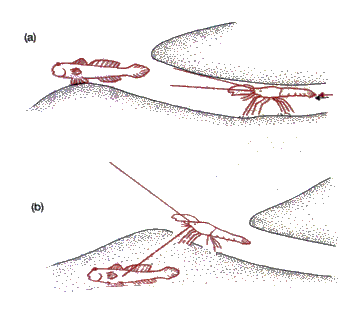
Los langostinos del género Alpheus excavan
madrigueras, y el pez gobio (Cryptocentrus) las utiliza como lugares
seguros en un ambiente que proporciona poca o ninguna protección. El langostino
es casi completamente ciego, y cuando abandona su escondrijo mantiene sus
antenas en contacto con el pez, que le avisa de cualquier perturbación (fig.
13.2). El gobi obtiene un lugar para vivir en un ambiente de sedimentos con
abundante comida. Por su lado, el langostino se beneficia con la adquisición de
un sistema de alarma óptico que le permite abandonar con seguridad su
escondrijo durante períodos cortos para buscar su alimento en los sedimentos
del exterior (Fricke, 1975). 13.2.3 El pez payaso y las anémonas Entre los habitantes de los arrecifes tropicales de coral (en donde las propias corales son mutualistas — véase más adelante) se encuentran diversas pautas mutualistas de comportamiento. El pez payaso (Amphiprion) vive cerca de una anémona marina (por ejemplo, Physobranchia, Radianthus) y se esconde entre los tentáculos de la anémona tan pronto le amenaza un peligro. Mientras se halla dentro de la anémona, el pez obtiene de ella un recubrimiento mucoso que le protege contra los nematocistos urticante de la anémona (el mucílago de la anémona es una sustancia cuya función normal estriba en impedir la descarga de los nematocistos cuando entran en contacto tentáculos vecinos). Es evidente que el pez obtiene protección de esta relación, pero también la anémona sale beneficiada, ya que el pez payaso ataca a los demás peces que se acercan, incluidos los de aquellas especies que normalmente se alimentan de anémonas (Fricke, 1975). Continúa ...
Introducción Esta sección consta sólo de dos capítulos. Ambos tratan unos temas que, en principio, hubieran podido ser estudiados en capítulos anteriores del presente libro; pero, en ambos casos, la dilación tenía sus ventajas. En el capítulo 14 examinamos, y tratamos de comprender, la gama de ciclos vitales o «estrategias vitales» que muestran los organismos vivos; en el capítulo 15 examinamos, y tratamos de comprender, la gama de esquemas de abundancia que muestran los organismos vivos. Así pues, estos capítulos son, en cierto sentido, sendos resúmenes de los dos niveles de la jerarquía ecológica que hemos estudiado hasta el momento: el capítulo 14 trata de los organismos aislados, y el capítulo 15 de las poblaciones. Los dos capítulos han sido colocados en esta parte del libro porque necesitaremos aplicar la información de los capítulos precedentes con el fin de entender los temas estudiados en la presente sección. En el capítulo 14 demostraremos que el ciclo vital de un organismo puede ser considerado a menudo como un reflejo del ambiente en que vive — pero este ambiente está formado, en gran medida, por los otros muchos organismos con los que entra en interacción. Análogamente, la abundancia de un organismo es, en esencia, un reflejo de los efectos combinados de las numerosas interacciones en las que toma parte. Así pues, si deseamos entender estos efectos, debemos primero llegar a comprenderlos por separado. Por consiguiente, las parte 1 y 2 del libro pueden ser consideradas como una preparación necesaria para los capítulos 14 y 15. A pesar de este razonamiento, puede parecer una lástima que la presente sección interrumpa el flujo aparentemente lógico que va desde el individuo hasta la población, y desde la interacción a la comunidad. Pero, mirándolo de otro modo, podríamos pensar que es extraño que esta sea la única interrupción de la secuencia. Cada aspecto de la ecología sólo puede ser entendido totalmente si se hace referencia a todos los demás aspectos — superiores, inferiores e iguales en la jerarquía. Por citar sólo un ejemplo: al principio del libro hemos hablado de las respuestas de los individuos a las condiciones, y estas respuestas constituyeron luego el telón de fondo para las interacciones de los organismos con otros organismos, y para la construcción de las comunidades de las que forman parte. Pero la respuesta de un organismo a las condiciones ambientales se halla influida a su vez, en gran medida, por la presencia de otros organismos que interaccionan con él, y por la estructura física y biológica de la comunidad circundante. Por consiguiente, la estructura «lógica» del libro es artificial. Ha sido impuesta por nosotros, para mayor comodidad, sobre una ciencia que quedaría mejor descrita en forma de disposición multidimensional de temas, con flechas que indicaran las influencias y que fluyeran en todas direcciones. En esta sección tomamos algunas flechas que van en dirección opuesta a la que hemos seguido a lo largo de la jerarquía.
Introducción En la Naturaleza, las áreas de tierra y los volúmenes de agua contienen agrupaciones de diferentes especies, en diferentes proporciones y que hacen diferentes cosas. Estas comunidades de organismos tienen unas propiedades que son la suma de las propiedades de los individuos más sus interacciones. Son estas interacciones las que hacen que la comunidad sea más que la suma de sus partes. Del mismo modo que parece razonable que un fisiólogo estudie el comportamiento de diferentes tipos de células y tejidos, e intente luego utilizar lo que sabe sobre sus interacciones para explicar el comportamiento del organismo entero, el ecólogo puede utilizar sus conocimientos sobre las interacciones entre los organismos para intentar explicar el comportamiento y la estructura de la comunidad entera. De hecho, cabe esperar que surjan esquemas estructurales en cualquier entidad formada por componentes que interaccionan entre sí, ya sean vivos o inertes. Por ejemplo, cuando se vierten cristales de ferrocianuro potásico en una solución de sulfato de cobre, se forma un bosque de estructuras parecidas a ramas que consume recursos (iones de cobre y de ferrocianuro) y que desarrolla una estructura predecible. La ecología de las comunidades es el estudio de las propiedades que se observan en la estructura y el comportamiento de las agrupaciones biológicas con múltiples especies. Estudiamos primero la naturaleza de la comunidad, es decir, lo que entendemos por comunidad. Por lo general nos referimos a alguna unidad del mundo natural que nosotros (investigadores humanos) podemos catalogar de acuerdo con unos rasgos que significan algo para nosotros. Imponemos un proceso antropocéntrico de selección al decidir lo que consideraremos una comunidad. Es muy importante darse cuenta de que las categorías establecidas pueden carecer de toda relevancia para las vidas de los organismos individuales de las comunidades. Nosotros podemos reconocer como comunidad a un estuario, o al rumen de una vaca, o a un roble, pero es posible que estos mismos elementos sean bastante irrelevantes para un langostino, un protozoo del rumen o una oruga. Para una oruga del roble, la única comunidad que tiene importancia para su vida consistirá probablemente en unas pocas hojas de una rama, junto con algunos competidores y depredadores que visitan su grupo de hojas. La visión de la comunidad que tiene cada organismo diferirá de una especie a otra. En el capítulo 16 imponemos nuestra perspectiva humana y describimos una serie de esquemas de la composición de las comunidades en el espacio y el tiempo. En el capítulo 17 examinamos los modos en que los organismos y sus alimentos se encuentran en la comunidad dentro de una red de elementos con interacción, dentro de la cual se mueve la energía y la materia. Más tarde (capítulo 21) tratamos de nuevo este tema y consideramos las implicaciones de la estructura de la red trófica sobre la dinámica de las comunidades y, en particular, aquellos aspectos de la estructura que contribuyen a la estabilidad. Los capítulos 18 y 19 tratan de nuevo unos temas mencionados ya en capítulos anteriores y examinan el grado en que la competencia, la depredación y la perturbación influyen sobre los esquemas que reconocemos a nivel de la comunidad. El capítulo 20 está dedicado a un área de la ecología que ha resultado extraordinariamente fructífero en la producción de nuevas perspectivas ecológicas —las islas y las comunidades insulares. Finalmente, intentamos realizar una síntesis (capítulo 22). La ciencia de la ecología es joven, y en ella sólo está empezando a surgir una síntesis. Utilizando de nuevo una analogía que ya hemos empleado antes, podernos decir que el estudio de la ecología al nivel de la comunidad se parece un poco a realizar un estudio sobre los relojes. Se puede reunir una colección de ejemplares y clasificar el contenido de cada uno de ellos. Podemos reconocer las características que tienen en común en cuanto al modo en que están construidos y a la función que desempeñan. Podemos reconocer esquemas y jerarquías dentro del conjunto de la colección. Pero para comprender como funcionan los relojes, será necesario desmontarlos, estudiar sus piezas, y volver a ensamblarlas. Habremos comprendido la naturaleza de las comunidades naturales cuando hayamos podido estudiar todas sus piezas y sepamos cómo volver a montarlas.
La información ofrecida está sujeta a cambios del Editor y/o el Vendedor del libro. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||